![]()
Habitat preservation or transplantation: which is better for Hong Kong seagrass conservation?
Habitat preservation or transplantation: which is better for Hong Kong seagrass conservation?
by Terence Ching-wai FongSeagrass beds are regarded as important habitats in Hong Kong and ecological assessment will be needed if a proposed development will affect an established seagrass bed of any size (Environmental Impact Assessment Ordinance). Despite the recognized importance of seagrasses, declines in seagrass meadows are global (Walker and McComb, 1992). These declines may result from natural events such as 'wasting disease' (Den Hartog, 1987) or high energy storms (Patriquin, 1972). Most seagrass loss, however, has resulted from human activities such as eutrophication (Bulthuis and Woelkerling, 1983; Cambridge and McComb, 1984; Neverauskas, 1987), sedimentation (Kirkman, 1978; Bulthuis et al., 1984; Lee, 1997), land reclamation and changes in land use (Kemp et al., 1983).
The rapid development experienced in recent decades has caused a decline in seagrass areas in Hong Kong. There is no information on the size and distribution of Hong Kong seagrass beds before the 1970s. Shatin, Tai Po and Plover Cove had large areas of mangroves and are believed to have had large seagrass beds in the past. Following reclamation and reservoir construction, all the coastal shores including those mangroves and seagrasses were destroyed. In addition, many small bays were completely dammed to form fishponds in the past, which may also have limited the abundance and distribution of seagrasses. The largest Zostera japonica bed in Lai Chi Wo is just located just beside such an impoundment. Halophila ovata was previously recorded in Deep Bay, Tai Tam and Hoi Sing Wan (Starfish Bay), but is now gone. It is difficult to estimate how much seagrass has been destroyed because of the lack of baseline information. However, there is some good evidence of serious decline in the total area of seagrass beds in Hong Kong in the past 20 years. Four recent case studies of conflicts between developments and seagrasses are discussed below.
Case 1: Residential development at Seung Sze Wan, Sai Kung
Zostera japonica and Halophila ovata were newly found in Seung Sze Wan, Sai Kung last year. Within one year, the H. ovata expanded from several small patches covering 10 m2 to about 1 hectare large now (recent survey on 14 June 1999). In addition to the original patch of Z. japonica found in 1998, several small patches were newly formed in the lower intertidal region. The total area of Z. japonica at Seung Sze Wan is now 20 m2. Due to the lack off background information, an old villager of Seung Sze Wan was interviewed. The old villager said that seagrasses were commonly found at Seung Sze Wan in the 1970s. Most seagrasses disappeared when building of residential houses started and the water quality deteriorated. The large patches of seagrasses reoccurred only in recent years after completion of the construction. The recent seagrass beds may derive from the seed bank of the sand flat or represent recolonization from other seagrass populations. The survival of seagrasses is therefore mainly influenced by anthropogenic effects. If allowed to grow in suitable habitats naturally without any artificial disturbance, they can recover and survive very well.Case 2: Development of Chek Lap Kok international airport
The development of the Chek Lap Kok new international airport has threatened the survival of nearby seagrasses. Reclamation and construction works there increased the sediment loading of the seawater and hence threatened the populations of Zostera japonica and Halophila ovata at San Tau, Lantau (Lee 1994; 1997). Z japonica and H. ovata, fortunately, are still surviving at this site, but most plants died during the peak period of reclamation. The population of Z. japonica recovered gradually after the completion of the Chek Lap Kok international airport (Fong, 1998a). After a recent survey on 11 June 1999, however, it is apparent that the native Z. japonica at San Tau has nearly totally disappeared; only several small colonies with a total area of less than 0.04 m2 remain. Only one of the five transplanted patches of Z. japonica (from Lai Chi Wo) remains, and has enlarged from 0.06 m2 to 15 m2. Under the highly turbid water, the health of the perennial plant Z. japonica was very weak. Once the construction was completed and the water quality improved, Z. japonica could start to recover. Heavy algal blooms have occurred every year since the completion of the airport, however, further weakening the seagrass. Heavy epiphytic growth seriously affects the above-ground component of Z. japonica by increasing the chance of sloughing weakened leaves through wave surge (Fong, I 998b). This, combined with the shading effect of algal blooms, has led to death of the seagrass. The relatively good health of transplanted patches has meant that some of them can survive and expand. As Halophila ovata is an annual and opportunistic plant, it recolonized San Tau and extended to Tung Chung Bay after a short period of time. Unlike Zostera japonica, H. ovata can survive under low light intensity, and is usually found in the lower intertidal zone. San Tau and Tung Chung are suitable habitats for H. ovata, and as the anthropogenic stresses were reduced, many patches ranging from 3 m to 10 m diameter were found along the coasts.Further development and reclamation in Tung Chung would impose further hazards on the seagrasses. Most of the coastline of Tung Chung Bay will be reclaimed, leaving a homogeneous concrete wall and no more suitable habitats for seagrasses. In addition, the increasing human population at Tung Chung New Town, close to San Tau, may further threaten the seagrass populations through sewage discharge and increased disturbance. The elevated nutrient loading of seawater resulting from increasing housing development is a common cause of seagrass decline (Short et al. 1996). Algal blooms could occur continuously, causing further decline of Z. japonica in San Tau. Further development and reclamation may also change the hydrology of Tung Chung Bay, possibly resulting in increased coastal erosion, which will wash out sediments and change the substratum of the seagrass bed. Therefore seagrasses would no longer survive in San Tau even though the area was designated as an SSSI.
Case 3: Bridge joining Shekou to Sheung Pak Nai
In order to improve vehicular transportation, the Governments of the Hong Kong Special Administrative Region and the People's Republic of China will build a bridge joining Shekou to Sheung Pak Nai, and the cross-link is expected to be completed by 2005 at the earliest (SCMP, 10/6/99). The construction of the bridge, like the development of the new airport, may seriously jeopardize the survival of seagrass, Halophila beccarii, which inhabits the southern coastline of Deep Bay. Deep Bay is already heavily polluted and conditions may worsen as the populations of the northwest New Territories of Hong Kong and the Shenzhen Special Economic Zone increase further. Reclamation and dredging will form a major component of the bridge construction programme and intertidal mud/sand flats may also be destroyed by the construction of the associated transportation network. Such development work in conjunction with pollution will probably severely impact the survival of H. beccarii. The H. beccarii beds in Deep Bay are well known nursery grounds for horseshoe crabs. Some sand patches and freshwater streams along the shoreline of Deep Bay are the breeding and spawning sites of the horseshoe crabs Carcinoscorpius rotundicauda and Tachypleus tridentatus (Huang et al., 1998). Such bridge construction seems unavoidable due to economic concerns and the involvement of the mainland government. Even though the turbidity of seawater in Deep Bay is naturally high, H. beccarii will still not be immune from the additional sedimentation. To minimize impacts on seagrass beds, the sedimentation caused by the associated constructions should be controlled. The construction of the transportation network should be sited inland, not close to the shore. Most important is to preserve and protect coastal shores, otherwise all the habitats suitable for seagrasses and horseshoe crabs will be destroyed.Case 4: Route connecting Mui Wo and Tai Ho
A new route connecting Mui Wo and Tai Ho to the Lantau Expressway was proposed and generated lots of argument recently. It is another example of contradiction between conservation and development. As most ecological concern and studies focused on the freshwater streams and the freshwater fishes, a preliminary coastal survey was carried out by myself on 11 June 1999. Halophila beccarii beds covering about 500 m2 total area were found in Tai Ho Wan. Many juveniles and a mating pair of horseshoe crabs were also found in the seagrass bed and adjacent mudflat. Construction of the new route not only impacts on the natural freshwater stream and associated flora and fauna, but also the seagrass H. beccarii and horseshoe crabs through either habitat destruction or sedimentation.
Creatures have their own way to struggle for life, Zostera japonica and Halophila ovata in Seung Sze Wan and San Tau are good examples. Both species, especially H. ovata, can recolonize damaged habitats once the anthropogenic stresses are removed and there is a return to favourable conditions. Development may not always result in local seagrass extinction, but if the habitats are totally destroyed, e.g. reclaimed for other uses or converted to a concrete wall, seagrass would be sacrificed. Restoration seems to be one remedy for declines in seagrass beds, but this is an inherently complicated process and has not yet been demonstrated to consistently prevent net habitat loss (Fonseca, 1989; Fonseca et al., 1994). Moreover, unless seagrasses are established over reasonably large areas, sediment movement and grazing may lead to an unsuccessful restoration. It is often difficult to find a site to fulfill the biological and physical requirements for persistence of restored seagrasses. Hong Kong is a small area, and most natural low-gradient coastlines have been or are being destroyed. Remaining suitable areas for rehabilitation or restoration of seagrasses are limited. If seagrass transplantation or restoration needs to be carried out because of a total destruction of existing seagrass habitats, I am afraid there will be no more suitable sites or potential areas for them to survive in. Even if seagrass beds can be restored, preservation is still the most cost-effective way to sustain seagrass resources. If developments or constructions just partly damage seagrass beds, without destroying the whole habitat, seagrasses can recover later under suitable management. The recovery duration depends on the degree of impacts and duration of the construction. Restoration, therefore, should be the last resort for unavoidable developments that endanger the survival of seagrasses. Transplantation should be performed only after possibilities for minimizing and mitigating damage to existing seagrass beds have been explored. The latter can be achieved through measures such as consideration of alternative sites for development, and design modifications to reduce detrimental effects upon seagrasses during and after construction. These are especially important for the developments in Deep Bay and Tai Ho.
Bulthuis, D.A. and Woelkerling, W.J. 1983. Biomass accumulation and shading effects of epiphytes on the leaves of the seagrass, Heterozostera tasmanica in Victoria, Australia. Aquatic Botany 16: 137-148.
Bulthuis, D.A., Brand, G.W. and Mobley. M.L. 1984. Suspended sediments and nutrients in water ebbing in from seagrass-covered and denuded tidal mudflats in a southern Australian embayment. Aquatic Botany 20: 257-266.
Cambridge, ML and McComb. A.J. 1984. The loss of seagrasses in Cockburn Sound, Western Australia. I. The time course and magnitude of seagrass decline in relation to industrial development. Aquatic Botany 20: 229-243.
Den Hartog. C. 1987. "Wasting disease" and other dynamic phenomena in Zostera beds. Aquatic Botany 27: 3-14.
Fong T.C.W. 1998a. Distribution of Hong Kong seagrasses. Porcupine! December 1998, Number 18.
Fong, C.W. 1998b. Some aspects of ecology of seagrass Zostera japonica in Hong Kong. 202pp. M. Phil. Thesis, Department of Ecology and Biodiversity, University of Hong Kong, Hong Kong.
Fonseca, M.S. 1989. Sediment stabilization by Halophila decipiens in comparison to other seagrasses. Estuarine, Coastal and Shelf Science 29: 501-507.
Fonseca, MS, Kenworthy, W.J., Courtney, F.X. and Hall, M.O. 1994. Seagrass planting in the southeastern United States: methods for accelerating habitat development. Restoration Ecology 2:198-212.
Huang. Q., Chiu, H. and Morton, B. 1998. Nursery beaches for horseshoe crabs in Hong Kong. Porcupine! December 1998, Number 18, pp. 9-10.
Kemp, W.M., Boynton, W.R., Twiggy, R.R., Stevenson, C. and Means, J.C. 1983. The decline of submerged vascular plants in Upper Chesapeake Bay: Summary of results concerning possible causes. Marine Technology Society. Journal 17: 78-89.
Kirkman, H. 1978. Decline of seagrass in northern areas of Moreton Bay, Queensland. Aquatic Botany 5: 63-76.
Lee, S.Y. 1994. Seagrass Rehabilitation at Tung Chung: Final Report. Report to the Hong Kong Government. 71 pp.
Lee, S.Y. 1997. Annual cycle of biomass of a threatened population of the internal seagrass Zostera japonica in Hong Kong. Marine Biology 129: 183-193.
Neverauskas, V.P. 1987. Monitoring seagrass beds around a sewage sludge outfall in South Australia. Marine Pollution Bulletin 18: 158-164.
Patriquin, D.G. 1972. Carbonate mud production by epibionts on Thalassia: an estimate based on leaf growth rate data. Journal of Sedimentary Petrology 42: 687-689.
Short. F.T., Burdick, D.M., Granger, S. and Nixon, S.W. 1996. Long-term decline in eelgrass, Zostera marina L., linked to increased housing development. In Seagrass Biology: Proceedings of an International Workshop. Rottnest Island. Western Australia. 1996. Kuo, J., Phillips, R.C., Walker, D.I. and Kirkman, H. (eds.), 291-298. Sciences, UWA.
Walker, D.I. and McComb, A.J. 1992. Seagrass degradation in Australian coastal waters. Marine Pollution Bulletin 25: 191-195.
A new genus of mycoheterotrophic flowering plants
by Richard Saunders
The Corsiaceae are a small family of mycoheterotrophic herbs, related to the Thismiaceae and Burmanniaceae. Only two genera have previously been described: Corsia, with ca. 26 species, restricted to New Guinea, Australia and the Solomon Islands; and Arachnitis, with two species from South America. An unusual herbarium specimen collected in Guangdong has been discovered that evidently belongs to the Corsiaceae but cannot be accommodated in either of the existing genera; a new genus and species, Corsiopsis chinensis, has accordingly been described (D.X. Zhang, R.M.K. Saunders & C.-M. Hu, Systematic Botany 24 (1999) 311-314). This is important since it represents the first record of the family in Asia and the northern hemisphere.
The flowers of Corsiopsis are solitary, epigynous and highly zygomorphic, with the six tepals in two distinct whorls. The posterior sepal (or labellum) is larger and distinct from the other tepals: it is bladder-like, remaining as a hood over the flower, with the margin proximally imbricate. The five other tepals (consisting of two sepals and three petals) are long and tentacle-like, reaching ground level. Similar elongated tepals have been reported for several species of Thismia; they presumably perform a similar function in Corsiopsis chinensis, providing access routes for insects of the forest floor. The bladder-like posterior sepal possibly acts as a "trap", so that the insects have difficulty leaving the flower, and consequently increase the chance of successful pollination.
Floral sexuality in the Corsiaceae has been the source of some confusion. The flowers of Corsia are hermaphroditic, although various authors have described them as protandrous, noting that the stigmas are poorly developed in young flowers with fully developed anthers, and that the anthers abscise when the stigmas become fertile. Arachnitis has hermaphroditic flowers that show extreme protandry: the stamens abscise during the female stage of development, leaving only small remnants of the filaments, which are often misinterpreted as staminodes. Although Corsia and Arachnitis possess bisexual flowers, Corsiopsis flowers appear to be truly unisexual. It is significant that there is no evidence of immature ovaries in male flowers or abscised stamens in female flowers of Corsiopsis; the reported unisexuality is therefore not believed to be associated with differential floral organ development or abscission. The stamens of Corsiopsis are not connate, and possess a distinctive obtuse apical extension of the connective. There are three connate and sessile stigmas in female flowers.
The geographical distribution of the Corsiaceae is markedly disjunct, occurring in South America (Arachnitis), Australasia (Corsia) and China (Corsiopsis). South-American-Australasian disjunctions are not uncommon in flowering plants, and result from a Gondwanan tectonic link through Antarctica. The South American-Australasian-Chinese disjunction is more unusual although similar patterns are observed at both the familial (Stylidiaceae) and generic (Coriaria) levels. Highly disjunct distribution patterns are relatively common in mycoheterotrophic flowering plants, although this may be an artefact of the rarity of collections due to their ephemeral emergence during seasonal rains, and their small size and inconspicuous coloration.
P.14-15
The conservation status of Cyperaceae in Hong Kong
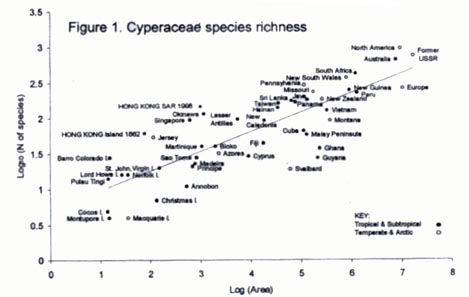
by Julia ShawHong Kong's biodiversity is fundamentally interesting as species numbers are higher than comparable areas elsewhere (Corlett & Turner, 1997; Shaw, in prep.; Fig. 1). The most reasonable explanations for this are Hong Kong's biogeographical location, a wide range of local habitats and a history of relatively intense plant collection (e.g. Bentham, 1861; Dunn & Tutcher, 1912; Anon., 1994).
Until the last millennium, East Asian forest was, uniquely, continuous from the equator to the Arctic (Kira, 1995). The establishment of open habitat as a result of human impact has added to the wide range of Hong Kong habitats from wetlands to high altitude forests (Ashworth et al., 1993; Dudgeon & Corlett, 1994; Shaw, in prep.). Unfortunately, wetlands and lowland forests are under threat from development to cater for predicted population growth of the 21st century. Furthermore, lowland habitats are generally outside of the Country Park system (Chu, 1998).
Status by habitat
The Cyperaceae (reeds, rushes and sedges) is the fifth largest plant family in Hong Kong (Anon., 1993) with representatives in all major terrestrial habitats (Shaw, in prep.). A recent two year field survey and review of various herbaria (e.g. Kew, Edinburgh, South China Institute of Botany, Hong Kong AFD, Chinese University of Hong Kong and the University of Hong Kong) has generated a list of 137 species (Shaw, in prep.). Wetlands are the centre of Cyperaceae species richness (75 species) while forest habitats are the centre of Cyperaceae species endemism (3 species). At least 50% of Hong Kong Cyperaceae are, therefore, threatened by development of Hong Kong's lowland habitats.Status by range
The Cyperaceae is not listed in the China Plant Red Data Book, while 10 Hong Kong angiosperms out of 388 taxa are (IUCN, 1978; Lai et al, 1999). Locally and globally rare Hong Kong Cyperaceae species were identified from the recent field survey and review of herbaria, literature and the Internet, with feedback from specialists worldwide (Shaw, in prep.). A comparison of the distribution data for each species was made using WORLDMAP rapid assessment software (Williams, 1996; Table 1, Table 2). The distribution data of six rare Hong Kong species recently collected by Ng Sai Chit has yet to be analysed. The recent collections are: Carex breviculmis R. Br., Carex filipes Franch. & Say. var. arisanensis (Hayata) Koy., Carex maculata Boott., Carex tristachya Thunb., Eleocharis chaetaria Roem. & Schult. and Schoenus falcatus R. Br.Management of Hong Kong Cyperaceae
There is currently no management of Hong Kong Cyperaceae, although protection is indirectly given to species growing within the Country Park system and Sites of Special Scientific Interest (SSSI) (Chu, 1998). The list above has been sent to the Agriculture and Fisheries Department of Hong Kong, according to the Technical Memorandum of the Environmental Impact Assessment Ordinance, and also to biodiversity information networks worldwide (Shaw, 1999a). There are 62 species of priority concern, 81 threatened species and 102 with important biogeographical populations.Seven wetland sites of rare Cyperaceae are highlighted for protection: Tai Wan, Po Toi (22°09.80'N 114°15.20'E); Tai Long Wan, East Sai Kung (22°24.50'N I 14°22.80'E); Lai Chi Wo, Wu Kai Sha (22°31.50'N 114°15.80'E); To Tei Wan (22°13.00'N 114°14.30'E); Siu Kau Yi Chau (22°17.30'N 114°03.00'E); and Cheung Lek Mei (22°25.00'N 114°11.70'E) (Shaw, in prep., 1999a). Further details are available from the recent Hong Kong Biodiversity Survey.
The only Hong Kong Cyperaceae species currently in cultivation is Gahnia tristis Nees at the Royal Botanic Gardens, Kew (Simpson, pers. comm.). Seed from the rare species above was limited and could not, therefore, be sent to the Royal Botanic Gardens seedbank. Advice is needed on appropriate in-situ and ex-situ conservation. Furthermore, molecular sequencing of Hong Kong Cyperaceae has yet to be undertaken, in particular for the rare genera Actinoschoenus, Diplacrum and Macherina (Muasya et al., 1998).
Table 1. Locally and globally rare Hong Kong Cyperaceae
Habitat Species Range Status details 4 Actinoschoenus filiformis Benth. 21-2-1 (NP) 1,2 Carex canina Dunn 1-0-1 (E) 1 Carex ligata Boott 2-0-1 1 Carex nexa Boott 2-0-1 6 Carex pumila Thunb. 31-0-1 (NP) 1 Carex speciosa Kunth 4-2-1 1 Carex tenebrosa Boott 1-0-2 (E) 6 Cyperus radians Nees Mey. 23-3-1 4,8 Diplacrum caricinum R.Br. 42-3-2 (NP) 3,4,8 Fimbristylis insignis Thwaites 26-0-1 (NP) 4,8 Fimbristylis longispica Steud. 19-3-1 (NP) 6 Fimbristylis sericea R.Br. 42-4-1 (NP) 1 Hypolytrum hainanensis (Merr.) Tang & Wang 2-0-1 (NP) 1 Machaerina ensigera (Hance) T. Koyama 1-0-1 (E) 1,2,3 Machaerina glomerata (Gaudich) T. Koyama 21-0-1 4,6,8 Schoenoplectus mucronatus (L.) Palla 60-4-1 (NP) 4,7 Schoenoplectus supinus (L.) Palla 55-0-1 (X) 2 Scleria corymbosa Roxb. 24-2-1 Key:
- One species is currently locally extinct (X), while three are regarded as endemic (E) and eight have not yet been collected in protected areas in Hong Kong (NP).
- Each species is prefixed by a code for Hong Kong habitats (1=dense forest, 2=semi-shaded forest, 3=shrubland, 4=wetland, 5=wasteland, 6=beach, 7=cultivated areas, 8=grassland); and
- suffixed by a distribution code for world range, South China status and Hong Kong status respectively.
- The world range (1-80) gives the number of WORLDMAP grid squares of 10 degrees latitude by 10 degrees longitude where the species has been recorded.
- The Hong Kong and south China status (1=rare, 2=restricted, 3=occasional, 4=frequent, and 5=common) is defined in Hong Kong by the number of population known (1=1-3sites, 2=3-6 sites, 4=10-20 sites and 5=more than 20 sites).
Table 2. Collection details of rare Hong Kong Cyperaceae
Species Collection details Actinoschoenus filiformis Benth. Hance, HK, 8. 1861, K!; SCNG 1354, Ma On Shan, 10.5.1997, HK Carex canina Dunn Dunn 7154, Lantau Peak 3.1911, K; SCNG 1354, Ma On Shan
10.5.1997, HK; SCNG 1171, Sunset Peak, Lantau, 5.3.98, HKCarex ligata Boott Wright, HK, 1853, K; Bentham 263, HK, 1858, K; SCNG 1334,
Pokfulam, HKCarex nexa Boott Bentham 790, HK, 1858, K; Rev. Faurie, Hance 15867, HK, 1898,
K; SCNG 1488, Nei Lak Shan, Lantau Peak, 10.8.1998; HKCarex pumila Thunb. Wright, HK, 1853; K; Shaw 393, Tai Long Wan, E. Sai Kung,
23.4.1995, HKCarex speciosa Kunth Dunn 5288, San Tau, Lantau, 16.6.1908, BM!; Shaw 252,
Ma On Shan, 19.10.1994, HK;SCNG 1333, Pokfulam, 23.4.1998Carex tenebrosa Boott Wilford 565, HK, 1853, K; Shaw 891, To Tei Wan, 19.3.1997, HK;
SCNG 1673, Wung Tung Chai, Tai Mo Shan, 24.12.1998, HK;
SCNG 1785, Sunset Peak, 11.3.1999, HKCyperus diffusus Vahl Hance 2149, HK, K; Hu SY 21961 Ng Tung Chai 2.1993, CHU Cyperus radians Nees Mey. Wright 565, HK, 1853, K Diplacrum caricinum R.Br. Hance 9932, HK, 11.1874, K; SCNG 896, Lai Chi Chong, Sai Kung,
2.11.1997, HK; SCNG 1504, Tai Tam, 8.9.1998, HKDiplacrum pygmaeopsis Kern Koy. SCNG 893, Lai Chi Wo, 2.11.1997, HK Fimbristylis insignis Thwaites (Hance 1871; Dunn & Tutcher 1912) Fimbristylis longispica Steud. Prof. Xing 9614; Shaw 645, Tai Po, 25.9.95, HK Fimbristylis sericea R.Br. SCNG 978, Siu Kau Yi Chau, HK!; SCNG 1002, HK Hypolytrum hainanensis Merr. Tang & Wang SCNG 1413, Tai Po Kau, 18.6.1998; SCNG 1779, Lion Rock, 7.3.1999,
HK SCNG, Cheung Lek Mei 1998, HKMachaerina ensigera Hance Koy. Hance, Pokfulam, 1.1883, K; Shaw 336, Shing Mun Country Park,
3.1995, HKMachaerina glomerata Gaudich Koy. Shaw 337, shing Mun Country Park, 3.1995, HK Schoenoplectus mucronatus L. Palla Bodinier 1324, Kowloon, 1895, E Schoenoplectus supinus L. Palla Wilford 271, HK, 1.1868, K Scleria corymbosa Roxb. Hance 2985, HK, 12.1858, BM; SCNG 1534, Sunset Peak, 1.10.1998 Monitoring is recommended for the genus Kyllinga (Shaw 1999b). K polyphylla Willd. has been recently introduced to Hong Kong (Corlett 1996) and is now running wild in grassland partly as a result of its robust rhizome system. It is burnt back from the banks of traditional fishponds in Luk Keng in the north-east of Hong Kong. Cyperaceae are not good invaders of native habitats (Daehler 1998), however, Kyllinga species have the potential to invade forest, Hong Kong's largest original habitat (Kira 1995). Kyllinga nemoralis (Forst.) Dandy, with distinct white inflorescence heads, currently grows in disturbed forest areas.
To maintain Hong Kong's biodiversity and habitats amidst the development pressures of the 21st century, in situ and ex situ management of the Cyperaceae is, therefore, a necessity.
Acknowledgements This work was funded by a studentship from the University of Hong Kong and facilitated by the hospitality of herbaria in the UK, Hong Kong and South China with feedback from specialists worldwide.
Anon (1994) Checklist of Hong Kong plants. Hong Kong herbarium, Agriculture and Fisheries Department Bulletin 1 (Revised): 87-90.
Ashworth, J. M., Corlett, R.T., Dudgeon, D., Melville, D. S. and Tang, W.S.M. (1993) Hong Kong Flora and Fauna: computing Conservation. Hong Kong Ecological Database. World Wide Fund for Nature, Hong Kong
Bentham, G. (1861) Flora Hongkonensis: a description of the flowering plants and ferns of the Island of Hong Kong. Lovell, Reeve, London.
Chu, W. H. (1998) Conservation of Terrestrial Biodiversity in Hong Kong. M. Phil. thesis, The University of Hong Kong.
Corlett, R. T. (1996) New sedge for Hong Kong. Porcupine! 14:4.
Corlett, R. T., Turner, I. M. (1997) Long-term survival in tropical forest remnants in Singapore and Hong Kong. In Tropical Forest Remnants: Ecology, Management, and Conservation of Fragmented Communities (ed. W. F Laurance, R. O. Bierregaard). The University of Chicago Press.
Daehler, C. C. (1998) The taxonomic distribution of invasive angiosperm plants; ecological insights and comparison to agricultural weeds. Biological Conservation 84: 167-180.
Dudgeon, D, Corlett, R. (1994) Hills and Streams: An Ecology of Hong Kong. Hong Kong University Press: 234.
Dunn S. T. and Tutcher, W. J. (1912) Flora of Kwantung and Hong Kong (China): An Account of the Flowering Plants, Ferns and Fern Allies together with Keys for their Determination. London: HMSO.
IUCN. (1978) The IUCN Plant Data Book. Gresham Press, Surrey.
Kira, T. (1995) Forest ecosystems of East and South-east Asia in a global perspective. Vegetation Science in Forestry. Kluwer Academic Publishers, Netherlands 1-21.
Lai, C.-c., Lau, S.-p., Cheung, K.-w, & Lam, Y.-w. (1999) Distribution and conservation of rare and endangered plants in Hong Kong. Journal of Tropical and Subtropical Botany 7(1):1-6.
Muasya, A.M., Simpson, D.A., Culham, A. and Chase, M. (1998). An assessment of suprageneric phylogeny in Cyperaceae using rbcL sequence data. Plant Systematic Evolution.
Shaw, J.C. (in prep.) Hong Kong Cyperaceae: The Taxonomy, Ecology & Geography. The University of Hong Kong (thesis in prep.).
Shaw, J.C. (1999a) Report on rare Hong Kong Cyperaceae. Unpublished report for the Agriculture and Fisheries Department of the Hong Kong S.A.R. Government.
Shaw, J.C. (1999b) Report to monitor the genus Kyllinga, a weedy genus with the potential to invade native habitats. Unpublished report for the Agriculture and Fisheries Department of the Hong Kong SAR Government.
Williams, P.H. (1996) WORLDMAP and prioritisation for conservation: integration of systematic data for conservation evaluation. Biodiversity assessment review: a guide to good practice London: HMSO 98.
P.16-18
Monster weed may threaten Hong Kong wetlands
By Richard CorlettNg Sai Chit has found a clump of the Giant Sensitive Plant, Mimosa pigra L., growing in wasteland at Tai Tung, near Three Fathoms Cove. The plant is mature and bears flowers and ripe, bristly pods. This species is unmistakable, since it is the only shrub in tropical East Asia which grows 3-5 m tall, has touch-sensitive bipinnate leaves, prickly stems and pink, globular flower-heads, 1-2 cm diameter. Oddly, Linnaeus considered it less sensitive than other species in the genus, hence the epithet "pigra", meaning lazy!
This Central American species has already become established over huge areas of the seasonal tropics and subtropics, in Southeast Asia, northern Australia and parts of Africa. It replaces species-rich wetlands with impenetrable, monospecific shrublands. In the Northern Territory of Australia it is considered the number one environmental weed and the "largest single threat" to wetlands. In northern Thailand I have seen it thriving in habitats, such as abandoned paddy fields, and a climate very similar to Hong Kong's. The only effective method of control so far is intensive, repetitive treatment with herbicides, although 11 biological control organisms have already been released in Australia.
I will ask AFD to destroy the plant(s) at Tai Tung and monitor the site for regeneration from the persistent seed bank. However, it is unlikely that this is the only site. If you see any more individuals, please report them to me and the AFD, and ensure that they are destroyed.
P.18
![]()
Back to Contents
Back to Porcupine Homepage
Go to Departmental Homepage